

 English
English French
French Spanish
Spanish Russian
Russian Korean
Korean Japanese
JapaneseWhat Is the Production Method of D Allulose Powder?
1 D-Allulose Overview
D Allulose is a hexose sugar and a C-3 epimer of D-fructose. Due to its high sweetness, low energy content, unique physiological functions and potential health benefits, D-allulose is considered to be a new sweetener with great potential. It has become a research hotspot in the field of rare sugar biosynthesis worldwide. However, D-allulose is extremely rare in nature and difficult to synthesize chemically. The biosynthetic method, on the other hand, has a simple procedure and has made great breakthroughs in recent years. Therefore, the author reviews the recent research progress in the biosynthesis of D-allulose, including the physicochemical properties, physiological functions, applications, in vivo metabolism, and enzymatic production of D-allulose, covering the sources, enzymatic properties, crystal structures, catalytic mechanisms, heterologous expression, and separation and purification processes of the key enzymes involved.
1.1 Physicochemical properties and sources of D-allulose
D-Allulose is a six-carbon sugar with a melting point of 96 °C, soluble in water, and a density of 1.35 g/cm3[1]. The sweetness of D-allulose is 70% that of sucrose[2]. D-Allulose is a reducing sugar and can participate in browning reactions during heat treatment. The calorific value of D-allulose was measured to be 0.029 kJ/g in a rat experiment, which is 0.3% of the energy of sucrose, indicating that D-allulose has almost zero energy [3-4]. D-allulose is very rare in nature, and plant sources are extremely rare [5]. Small amounts of allulose have also been found in some bacteria [6], and D-allulose is not found in animals. allulose has not been found in animals. However, D-allulose is also found in various foods, such as fruit juice that has undergone long-term heating treatment, and the content of D-allulose in various foods is closely related to the sugar concentration, temperature and heating time during the production process [2, 7].

1.2 Physiological functions and metabolism of D-allulose in the body
The physiological functions of D-allulose include: reducing the absorption of dietary D-fructose and D-glucose [8-9]; enhancing insulin resistance [7, 10-11]; anti-obesity activity [12-14]; and lowering blood lipids [15]. Studies on the metabolic pathway of D-allulose in rats have shown that: 1) the transport of D-allulose in the intestine is mediated by glucose transporter 5 , and has a lower affinity for D-fructose [16-18]; 2) D-allulose is not involved in glucose-related metabolism [19]; 3) D-allulose cannot be metabolized in animal livers, and thus cannot promote liver energy production [20]. Approximately 98% of D-allulose is excreted from the body in the form of urine and feces after oral or intravenous administration, and only a small amount of D-allulose is broken down into short-chain fatty acids by the action of microorganisms in the cecum [21].
1.3 Applications of D-allulose
D-Allulose was approved as a Generally Regarded As Safe (GRAS) product by the US FDA in 2011 and is permitted as a food ingredient and dietary supplement [4, 7]. Studies have shown that the maximum intake of D-allulose is 0.55 g per kg of body weight per day, and that it will not cause diarrhea in humans within this range [8-9].
D-Allulose has broad market potential in the food industry due to its low calorie content, high sweetness and strong reducing properties. For example, D-allulose can comprehensively improve the properties of egg white protein through the Maillard reaction, such as excellent gel strength, emulsifying stability, foaming properties and antioxidant activity [22-23]. D-allulose can also improve the quality of fermented milk products, regulating the strong sour taste of yogurt caused by overfermentation, but does not affect the probiotic activity of the fermentation strain and the probiotic health benefits it imparts to the fermented product [ 21]. In addition, the use of 25% D-allulose in baking, in combination with other additives, can produce sugar-free cakes [24].
D-Allulose also has broad application potential in other fields. For example, plant-based materials made from D-allulose can be used as permanent, waterproof, environmentally friendly, light-transmitting films for optical devices and liquid crystal displays [25]. D-Allulose is also the first sugar-based insect repellent to be discovered, and has some positive effects in inhibiting the growth of parasites [26-27]. D-allulose is also a precursor to other hexoses and plays an extremely important role in the production of D-allose [28-29] and D-allitol [30] plays an extremely important role.
2.Biological enzyme method
The chemical synthesis method of D-allulose has common disadvantages that are difficult to overcome, such as difficulties in isolation, many by-products, and the generation of chemical waste. Therefore, the green and environmentally friendly biological enzyme production method has gradually received widespread attention worldwide.
2.1.1 Source of DTEase family enzymes
The D-tagatose 3-epimerase family (DTEases) is an enzyme that catalyzes the isomerization of keto monosaccharides at the C3 position and is also the core enzyme for the production of rare sugars [32]. The DTEase family of enzymes includes: D-tagatose 3-epimerase (DTEases) [33], D-allulose 3 -epimerase (DAEase) [35], ketose 3-epimerase [36], all of which have the common property of catalyzing the conversion of D-fructose to D-allulose.
The gene encoding DTEase has been sequentially isolated from Clostridium cellulolyticum H10[37], Ruminococcus sp.[38], Clostridium scindens[34] and Desmospora sp.[35], but there is still a need to explore more sources and more efficient DTEase[32, 37, 39-40] for the industrial production of D-allulose.
2.1.2 Enzyme properties of DTEase
Among the DTEase family of enzymes, D-allulose-3-epimerase (DTEase) has the highest efficiency in catalyzing the reaction of D-fructose to D-allulose. The catalytic efficiency (Kcat/Km) of DAEases from C. cellulolyticum and A. tumefaciens is 186.4 and 205 L/(mmol·min), respectively, which is higher than that of DTEase (D-tagatose 3-epimerase) from Clostridium sp. (141.4 L/(mmol·min)) and Ruminococcus sp. (51 L/(mmol·min)) [34, 37–38], see Table 1.
Metal ions are fixed by binding to D-fructose and play a crucial role in the conversion of D-fructose to D-allulose. The Asp183 and His209 residues effectively bind the substrate through the metal ions. The DTEase family of enzymes has significantly different degrees of dependence on metal ions Mn2+, Co2+ and Mg2+ [41, 44- 45]. However, the enzymatic activity of DAEase from C. cellulolyticum and DTEase from C. scindens is strictly dependent on the metal ions Mn2+ and Co2+[35, 37].
2.1.3 Crystal structure of DTEase
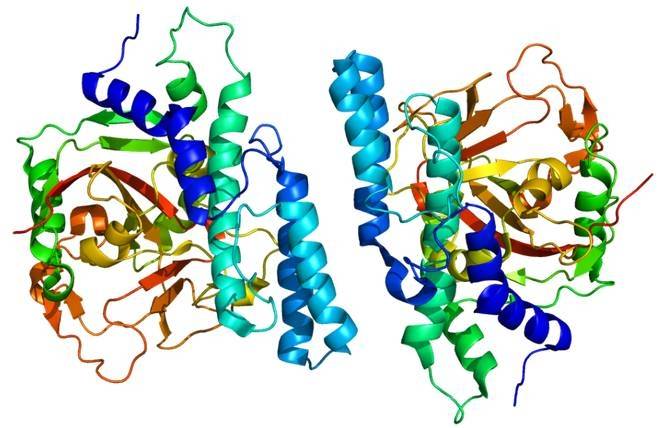
Among the DTEase family of enzymes, the DAEases from A. tumefaciens and C. cellulolyticum exhibit the closest form of tetramer. In the DAEase tetramer, 34 hydrogen bonds are formed between amino acid residues in the two subunits. The high catalytic activity of DAEase is attributed to the wide interfacial solvent-accessible area, which is caused by extensive interactions between the two dimers of the DTEase family enzyme [42]. In addition, DAEase expressed by recombinant strains still exhibits excellent enzymatic properties. The heterologous expression of DAEase from C. cellulolyticum H10 has a longer half-life, higher kinetic parameters and higher thermal stability [37].
The quaternary arrangement of DAEase is an asymmetric unit consisting of four identical subunits A, B, C, and D. The active site contains four residues of the metal ion, octahedral coordination of two water molecules. These four subunits are dimers related by crystal symmetry, in which subunits A and D interact with each other, both in close contact with subunits B and C. The active site of DAEase is exposed on the same front side of the dimer. These stable dimers provide an excellent accessible surface for binding substrates on the front side of the dimer [16].
The hydrophobic groove of the active site and the accessible surface are located between the A and B subunits. The subunit side of DAEase is closed and exposed at both ends of the barrel structure. In the tetramer of DAEase, the two dimers are enclosed on the sides of the barrel structure. Each monomer (subunits A, B, C and D) is composed of 8 repeat units of (β/α) structures. Each monomer is composed of 13 α-helices and 8 β-folds, forming the main structure of the monomer [16].
2.1.4 Catalytic mechanism of DTEase
The catalytic action of the DTEase family of enzymes depends on the molecular arrangement of the subunits. Their active sites are located on the substrate to achieve efficient enzymatic reactions. Mn2+ and two water molecules and four amino acids (Glu, Asp, His and Glu) form an octahedral coordination, and these four amino acid residues are completely conserved in all ketose 3-epimerases. Six residues (Glul50/Glul52, Aspl83 / Aspl85, His209 / His211, Glu244 / Glu246, Glul56/ Glu158, His185/His188) from DAEase in A. tumefaciens and DTEase in P. cichorii are important for substrate binding and thermal stability in site-directed mutagenesis.
After the substrate replaces the water molecule, the active site undergoes a diastereoisomerization. Two residues, Glu50 and Glu244, in collaboration with Mn2+, remove the proton on C3 of D-fructose to form the cis-diol intermediate of D-allulose, and D-allulose is released from the position between the hydrogen bond and the water molecule on the DAEase active site.

At present, molecular modifications are being carried out on enzymes in the DTEase family to improve their catalytic activity and thermal stability. Site-directed mutagenesis was used to improve the thermal stability and catalytic behavior of L-rhamnose isomerase from Caldicellulosiruptor obsidiansis in the production of D-allulose. The hydrophobic residues in the β1-α1-loop were completely replaced with polar amino acids. Compared with the wild-type enzyme, the relative activities of the V48N/G5 9N/I63N and V48N/G59N/I63N/F335S mutants have relative activities that are 105.6% and 134.1% higher, respectively, than those of the wild-type enzyme [57]. Site-directed mutagenesis also improved the thermal stability of D-allulose-3-epimerase (DAEase) from Dorea sp. [50].
Research on the catalytic mechanism of DTEase family enzymes is still in its infancy, and the relationship between enzyme structure and catalytic function requires further in-depth study. In addition, the thermal stability and substrate specificity of DTEase still have some deficiencies. In the production of rare sugars, the substrate specificity of DTEase family enzymes should be further utilized to achieve the efficient and green production of functional rare sugars.
2.1.5 Heterologous expression of DTEase
Most DTEase family enzymes have been identified and isolated from bacteria, and the number of enzymes expressed in natural strains is far from meeting the requirements of applications. Therefore, constructing expression vectors and expressing them in heterologous organisms is of great significance in characterization studies and enzymatic applications.
Bacillus subtilis, Escherichia coli and yeast are commonly used to construct recombinant systems for DAEase expression. Unlike E. coli, Bacillus subtilis does not have an outer membrane, so the proteins it secretes can be released directly into the culture medium. Bacillus subtilis is also food grade and does not secrete heat-labile lipopolysaccharides (endotoxins) in the proteins it secretes. The engineered bacterium Escherichia coli (E. coli) has the advantages of a clear genetic background, a complete vector receptor system, rapid growth, simple cultivation, and recombinant stability. In addition, the yeast expression system has the characteristics of simple culture conditions, rapid growth, high expression levels, and simple operation. After translation, the protein can be processed and correctly modified. The disadvantages of the yeast expression system are low cloned gene expression, long fermentation time, incorrect protein glycosylation, and resistance to cell division. In addition, the high concentration of polysaccharides in the supernatant is not conducive to the purification of recombinant proteins.
Early researchers tended to use E. coli as the host bacterium to study the expression and enzymology of DTEase family enzymes [34, 37]. Recently, many researchers have used Bacillus subtilis and yeast as host bacteria to express DTEase family enzymes for the production of D-allulose [35]. The DAEase gene from A. tumefaciens was expressed in E. coli and K. marxianus after recombination and used for allulose production, and the yields of D-allulose reached 230 g/L [53] and 190 g/L [56] respectively. Comparatively, the DAEase enzyme expressed in A. tumefaciens using the E. coli expression system has the highest expression level to date, as shown in Table 2.
(1) E. coli expression system: The E. coli expression system has the advantages of low cost and high expression efficiency. The DTEase family of enzymes can be overexpressed in E. coli as soluble proteins. The recombinant DTEase enzyme is separated and purified using affinity chromatography, and the yield of D-allulose is 120–218 g/L, with a conversion rate of 24%–33% [34]. The yield and conversion efficiency of the DAEase enzyme expressed in recombinant Escherichia coli using the whole-cell response method from Agrobacterium tumefaciens were 230 g/L and 33%, respectively [53].
(2) Bacillus expression system: Recombinant Bacillus subtilis carrying the DAEase coding gene can overexpress DAEase enzyme in a high-efficiency and low-cost manner. Recombinant DAEase immobilized on an anion exchange resin matrix can promote the stable and efficient production of allulose. The recombinant DAEase expressed in Bacillus subtilis B. subtilis has an activity of 58.6 U/mg, which is higher than that in E. coli (8.95 U/mg). In addition, the regulatory element of DAEase expression also affects the amount and activity of the enzyme. In Bacillus subtilis B. subtilis, the vector pMA5- Pxy/A-RDPE can stably express DAEase with an enzyme activity of 95 U/mL. This value is higher than the enzyme activity expressed in E. coli E. coli with the pBluescript-SK-DTE vector [54].
3) Yeast expression system:
The exogenous DAEase gene can be highly expressed in recombinant S. cerevisiae [55] and Kluyveromyces marxianus [56]. The expression vector pRS424-TEFpr-ss-xy/A, which carries the DAEase gene from A. tumefaciens, can express a protein with a relative molecular mass of 33 000 in S. cerevisiae AN120 [55]. The xylose isomerase gene from T. thermophilus and the DAEase gene from A. tumefaciens are co-expressed in yeast spores to enhance synergistic catalysis. The two recombinant enzymes were immobilized and D-allulose was produced using D-glucose as the substrate [55]. The recombinant xylose isomerase catalyzes the conversion of D-glucose to D-fructose, and the recombinant DAEase converts D-fructose to D-allulose. Yang et al. provided a valuable approach to regenerate modified K. marxianus cells, which can produce D-allulose in a recycling catalytic manner [56]. The recombinant K. marxianus produced 190 g/L D-allulose from a substrate concentration of 750 g/L D-fructose within 12 h, and about 100 g of residual D-fructose was converted by the engineered bacteria into 34 g of ethanol. In addition, the idea of producing D-allulose by whole-cell biocatalysis has also been proposed [56].
2.2 Separation and purification of D-allulose
The separation and purification of D-allulose mainly includes the following two methods: The first method is ion exchange resin. Anion exchange resin matrix and simultaneous moving bed chromatography were used to immobilize DAEase enzyme to produce D-allulose from D-fructose as a substrate. Using ion exchange resin dialysis, R. sphaeroides SK011 cells can produce 6.5 g/L D-allulose from an initial substrate concentration of 50 g/L D-fructose [39], with a production rate of 0.82 g·h-1. For a mixed system containing D-allulose and D- fructose, D-allulose was first converted to gluconic acid and then purified to 91.2% by anion exchange resin [57].
The second method uses a biological method to purify D-allulose. In a mixed system containing D-allulose and D-fructose, D-allulose was obtained by using yeast to consume the remaining D-fructose and produce ethanol. In addition, a combination of osmotic evaporation, cation exchange chromatography and biological methods was used to separate and purify D-allulose, with a purity of up to 86.2% [58].
3 Discussion
D-Allulose is the C-3 epimer of D-fructose, which has many physiological functions and is widely used in food, medicine, and health care. After the US FDA passed a favorable measure in April 2019 to exclude allulose from the counting of added sugars and total sugars, major companies have begun to take a great interest in allulose. Currently, the world's major producers of D-allulose, including South Korea's CJ CheilJedang, the UK's Tate & Lyle and Japan's Matsutani Group, all use biological methods to produce D-allulose. Therefore, the construction of genetically engineered bacteria that can catalyze the production of D-allulose is an important foundation for the industrialization of D-allulose. Although allulose can currently only be legally used in a few countries, it is undoubtedly set to become a major sweetener in the future.
Although there have been major breakthroughs in the research of the biological enzyme method for D-allulose, in particular the rapid development of gene mining and cell construction technology, the industrial prospects for the biological production of D-allulose remain uncertain. Subsequent research can be carried out in the following two areas: 1) screening DTEase family enzymes with high activity and stability through gene mining methods, establishing efficient high-throughput screening methods, and molecularly modifying DTEase family enzymes to better meet the needs of industrialization, thereby achieving efficient conversion of D-fructose and D-allulose; 2) since the industrial preparation of D-allulose faces the problem of removal of residual D-fructose in the mixture after enzymatic catalysis, the separation and purification of D-allulose increases its production cost. At present, there are relatively few studies on the separation and purification and crystallization of D-allulose, indicating that downstream industrialization is relatively lagging behind. It is necessary to further reduce the industrial production cost of D-allulose and improve the production efficiency of D-allulose by simplifying downstream process steps, such as separation and purification, crystallization, drying and other process steps.
Figure 1 shows a more advanced green and recyclable process technology for D-allulose [16]. The entire reaction system is divided between bioreactor A (for sucrose hydrolysis and enzymatic conversion to D-allulose) and bioreactor B (for ethanol production, D-allulose separation and yeast propagation). Raw sugar cane juice or sweet sorghum juice is used as the raw material to produce sucrose. The engineered yeast used contains natural sucrose isomerase and a recombinant exogenous DTEase family enzyme gene, so the engineered yeast can produce D-glucose and D-fructose by hydrolyzing sucrose with sucrose isomerase. D-fructose is converted to D-allulose by the DTEase family enzyme at 55~60 °C, and the remaining D-glucose and D-fructose are fermented to produce ethanol at 27-30 °C, and the ethanol can be collected and used as fuel. The advantages of this method are low raw material costs, the use of intermediate products generated during the process as much as possible, the reduction of cumbersome separation and purification steps, as well as the reduction of waste formation, lower energy consumption and higher sugar yields.
4 Outlook
At present, D-allulose has been produced industrially at the tonne level in China, Japan, South Korea, the United States and the United Kingdom. To stand out in the fierce industry competition, enterprises producing D-allulose must improve the technology of each link in the process. At present, the main production barriers are the low activity of the isomerase, the low conversion rate and the low reuse frequency. Therefore, improving enzyme activity, stability and catalytic efficiency should be the key objectives of future research and development of the DTEase family of enzymes. The production cost of D-allulose can be reduced by improving the processes of decolorization, desalination, crystallization and drying. The rational design or modification of unreasonable processes using efficient high-throughput screening technology is a direct way to change the structure of the DTEase family of enzymes. By improving the DTEase family of enzymes and improving the production process, the production cost of D-allulose can be reduced and its price lowered, thus ensuring that D-allulose can be easily supplied to consumers.
Reference
[ 1 ] FUKADA K,ISHII T,TANAKA K,et al. Crystal structure,solubility,and mutarotation of the rare monosaccharide D-psicose[J]. Bulletin of the Chemical Society of Japan,2010,84(6):678-678.
[ 2 ] OSHIMAH,KIMURA I,IZUMORI K. Psicosecontents in various food products and its origin[J]. Food Science and Technology Research,2006,12(2):137-143.
[ 3 ] MATSUO T,SUZUKI H,HASHIGUCHI M,et al. D-psicose is arare sugar that provides no energy to growing rats[J]. Journal of Nutritional Science and Vitaminology,2002,48(1):77-80.
[ 4 ] MU W,ZHANG W,FENG Y,et al. Recent advances on applications and biotechnological production of D-psicose[J]. Applied Microbiology and Biotechnology,2012,94(6):1461-1467.
[ 5 ] Benjamin J Ayers,Jacqueline Hollinshead,Alexander W Saville,et al. Iteamine,the first alkaloid isolated from Iteavirginica L. in florescence[J]. Phytochemistry,2014,100(2):126-131.
[ 6 ] ZHANG L,MU W,JIANG B,et al. Characterization of D-tagatose-3-epimerase from Rhodobacter sphaeroides that converts D-fructose into D-psicose[J]. Biotechnology Letters,2009,31:857-862.
[ 7 ] ZHANG W,YU S,ZHANG T,et al. Recent advances in D-allulose:physiological functionalities,applications,and biological production[J]. Trends in Food Science and Technology,2016,54(54):127-137.
[ 8 ] HOSSAIN A,YAMAGUCHI F,HIROSE K,et al. Rare sugar D-psicose prevents progression and development of diabetes in T2DM model Otsuka Long-Evans Tokushima Fatty rats[J]. Drug Design,Development and Therapy,2015,9:525-535.
[ 9 ] HOSSAIN A,YAMAGUCHI F,MATSUO T,et al. Rare sugar D-allulose:potential role and therapeutic monitoring in maintaining obesity and type 2 diabetes mellitus[J]. Pharmacology & Therapeutics,2015,155:49-59.
[10] HOSSAIN M A,KITAGAKI S,NAKANO D,et al. Rare sugar D-psicose improves insulin sensitivity and glucose tolerance in type 2 diabetes Otsuka Long-Evans Tokushima Fatty(OLETF)rats[J]. Biochemical and Biophysical Research Communica- tions,2011,405(1):7-12.
[11] OCHIAI M,NAKANISHI Y,YAMADA T,et al. Inhibition by dietary D-psicose of body fat accumulation in adult rats fed a high-sucrose diet[J]. Bioscience Biotechnology & Biochemistry,2013,77(5):1123-1126.
[12] CHUNG Y M,LEE J H,KIM D Y,et al. Dietary D-psicose reduced visceral fat mass in high-fat diet-induced obese rats [J]. Journal of Food Science,2012,77(2):H53-H58.
[13] CHUNG MY,OH DK,LEE KW. Hypoglycemic health benefits of D-psicose[J]. Journal of Agricultural and Food Chemistry, 2012,60(4):863-869.
[14] IIDA T,YAMADA T,HAYASHI N,et al. Reduction of abdominal fat accumulation in rats by 8-week ingestion of a newly developed sweetener made from high fructose corn syrup[J]. Food Chemistry,2013,138(2-3):781-785.
[15] OCHIAI M,ONISHI K,YAMADA T,et al. D-psicose increases energy expenditure and decreases body fat accumulation in rats fed ahigh-sucrose diet[J]. International Journal of Food Sciences and Nutrition,2014,65(2):245-250.
[16] JIANG S,XIAO W,ZHU X,et al. Review on d-allulose:In vivo metabolism,catalytic mechanism,engineering strain construction,bio-production technology[J]. Frontiers in Bioengineering and Biotechnology,2020,8:26.
[17] RAN G,TAN D,ZHAO J,et al. Functionalized polyhydroxyalkanoatenano-beads as a stable biocatalyst for cost-effective production of the rare sugar D-allulose[J]. Bioresource Technology,2019,289:9-18.
[18] KISHIDA K,MARTINEZ G,IIDA T,et al. D-allulose is a substrate of glucose transporter type 5(GLUT5)in the small intestine [J]. Food Chemistry,2019,277:604-608.
[19] IWASAKI Y,SENDO M,DEZAKI K,et al. GLP-1 release and vagal afferent activation mediate the beneficial metabolic and chronotherapeutic effects of D-allulose[J]. Nature Communications,2018,9:17-25.
[20] MAENG H J,YOON J H,CHUN K M,et al. Metabolic stability of D-allulose in biorelevant media and hepatocytes:comparison with fructose and erythritol[J]. Foods,2019,8:13-18.
[21] KIMOTO-NIRA H,MORIYA N,HAYAKAWA S,et al. Effects of rare sugar D-allulose on acid production and probiotic activities of dairy lactic acid bacteria[J]. Journal of Dairy Science,2017,100(7):5936-5944.
[22] O'CHAROEN S,HAYAKAWA S,OGAWA M. Food properties of egg white protein modified by rare ketohexoses through maillard reaction[J]. International Journal of Food Science & Technology,2015,50(1):194-202.
[23] YAN Z,ZHANG H,GUAN Y,et al. Comparative study on the effects of D-psicose and d-fructose in the maillard reaction with β-lactoglobulin[J]. Food Science and Biotechnology,2013,22(2):341-346.
[24] LEE P,OH H,KIM S Y,et al. Effects of D-allulose as a sucrose substitute on the physicochemical,textural,and sensorial properties of pound cakes[J]. Journal of Food Processing and Preservation,2020,44(6),e14472.
[25] TAKEI S,HANABATA M. Eco-friendly,water-repellent,light-transparent film derived from psicose using nanoimprint lithography[J]. Materials Letters,2015,143:197-200.
[26] HARADA M,KONDO E,HAYASHI H,et al. D-allose and D-psicose reinforce the action of metronidazole on trichomonad[J]. Parasitology Research,2012,110(4):1565-1567.
[27] SATO M,KUROSE H,YAMASAKI T,et al. Potential anthelmintic:D-psicose inhibits motility,growth and reproductive maturity of L1 larvae of Caenorhabditis elegans[J]. Journal of Natural Medicines,2008,62(2):244-246.
[28] FENG Z,MU W,JIANG B. Characterization of ribose-5-phosphate isomerase converting D-psicose to d-allose from Thermotoga lettingae TMO[J]. Biotechnology Letters,2013,35:719-724.
[29] YEOM S J,SEO E S,KIM Y S,et al. Increased D-allose production by the R132E mutant of ribose-5-phosphate isomerase from Clostridium thermocellum[J]. Applied Microbiology and Biotechnology,2011,89(6):1859-1866.
[30] HAN W,ZHU Y,MEN Y,et al. Production of allitol from D-psicose by a novel isolated strain of Klebsiella oxytoca G4A4 [J]. Journal of Basic Microbiology,2014,54(10):1073-1079.
[31] SOENGAS R,IZUMORI K,SIMONE MI,et al. Kiliani reactions on ketoses:branched carbohydrate building blocks from d-tagatose and D-psicose[J]. Tetrahedron Letters,2005,46(34):5755-5759.
[32] JIA M,MU W,CHU F,et al. A D-psicose 3-epimerase with neutral pH optimum from Clostridium bolteae for D-psicoseproduction:cloning,expression,purification and characterization[J]. Applied Microbiology and Biotechnology,2014, 98(2):717-725.
[33] ZHANG L,MU W,JIANG B,et al. Characterization of D-tagatose-3-epimerase from Rhodobacter sphaeroides that converts D-fructose into D-psicose[J]. Biotechnology Letters,2009,31(6):857-862.
[34] MU W,ZHANG W,FANG D,et al.,Characterization of a D-psicose-producing enzyme,D-psicose 3-epimerase,from Clostridium sp.[J]. Biotechnology Letters,2013,35(9):1481-1486.
[35] ZHANG W,FANG D,XING Q,et al. Characterization of a novel metal-dependent D-psicose 3-epimerase from Clostridium scindens 35704[J]. Plos One,2013,8,e62987.
[36] YOSHIDA H,YOSHIHARA A,SUZUKI T,et al. Xray structure of a novel ketose 3-epimerase from Shinella zoogloeods which is capable of D-allulose production[J]. FEBS Open Biol,2019,9:257-265.
[37] MU W,CHU F,XING Q,et al. Cloning,expression,and characterization of a D-psicose 3-epimerase from Clostridium cellulolyticum H10[J]. Journal of Agricultural and Food Chemistry,2011,59(14):7785-7792.
[38] ZHU Y,MEN Y,BAI W,et al. Overexpression of D-psicose 3-epimerase from Ruminococcus sp. in Escherichia coli and its potential application in D-psicose production[J]. Biotechnology Letters,2012,34(10):1901-1906.
[39] ZHANG L,JIANG B,MU W,et al. Bioproduction of D-psicose using permeabilized cells of newly isolated Rhodobacter sphaeroides SK011[J]. Frontiers of Chemical Engineering in China,2009,3(4):393-398.
[40] PARK C,KIM T,HONG S,et al. D-allulose production from D-fructose by permeabilized recombinant cells of Corynebacterium glutamicum cells expressing D-allulose 3-epimerase Flavonifractor plautii[J]. Plos One,2016,11(7),e0160044.
[41] YANG J,TIAN C,ZHANG T,et al. Development of food-grade expression system for D-allulose 3-epimerase preparation with tandem isoenzyme genes in Corynebacterium glutamicum and its application in conversion of cane molasses to D-allulose [J].
Biotechnology and Bioengineering,2019,116(4):745-756.
[42] LI S,CHEN Z,ZHANG W,et al. Characterization of a D-tagatose 3-epimerase from Caballeronia fortuita and its application in rare sugar production[J]. International Journal of Biological Macromolecules,2019,138:536-545.
[43] ZHU Z,LI C,LIU X,et al. Biochemical characterization and biocatalytic application of a novel D-tagatose 3-epimerase from Sinorhizobium sp.[J]. RSC Advances,2019,9(6):2919-2927.
[44] ZHU Z,GAO D,LI C,et al. Redesign of a novel D-allulose 3-epimerase from Staphylococcus aureus for thermostability and efficient biocatalytic production of D-allulose[J]. Microbial Cell Factories,2019,18(1):1-10.
[45] TSENG W,CHEN C,HSU C,et al. Characterization of a recombinant D -allulose 3-epimerase from Agrobacterium sp. ATCC
31749 and identification of an important interfacial residue [J]. International Journal of Biological Macromolecules,2018, 112:767-774.
[46] YOSHIHARA A,KOZAKAI T,SHINTANI T,et al. Purification and characterization of D-allulose 3-epimerase derived from Arthrobacter globiformis M30,a GRAS microorganism[J]. Journal of Bioscience and Bioengineering,2017,123(2):170-176.
[47] HE W,JIANG B,MU W,et al. Production of D-allulose with D-psicose 3-epimerase expressed and displayed on the surface of Bacillus subtilis spores[J]. Journal of Agricultural and Food Chemistry,2016,64(38):7201-7207.
[48] ZHANG W,ZHANG T,JIANG B,et al. Biochemical characterization of a D-psicose 3 - epimerase from Treponema primitiaZAS-1 and its application on enzymatic production of D -psicose [J]. Journal of the Science of Food and Agriculture,2016,96(1): 49-56.
[49] ZHANG W,LI H,ZHANG T,et al. Characterization of a D-psicose 3-epimerase from Dorea sp. CAG317 with an acidic pH optimum and a high specific activity[J]. Journal of Molecular Catalysis B-Enzymatic,2015,120:68-74.
[50] ZHANG W,ZHANG Y,HUANG J,et al. Thermostability improvement of the d-Allulose 3-epimerase from Dorea sp. CAG317 by site-directed mutagenesis at the interface regions[J]. Journal of Agricultural and Food Chemistry,2018,66:5593-5601.
[51] PATEL S N,KAUSHAL G ,SINGH S. Characterization of a novel D-allulose 3-epimerase gene from the metagenome of a thermal aquatic habitat and D-allulose production by Bacillus subtilis whole-cell catalysis[J]. Applied and Environmental Mic- robiology,2019,1:1-8.
[52] CHEN Z,CHEN J,ZHANG W,et al. Improving thermostability and catalytic behavior of L-rhamnose isomerase from Caldicellulosiruptor obsidiansis OB47 toward D -allulose by site-directed mutagenesis [J]. Journal of Agricultural and Food Chemistry,2018,66(45):12017-12024.
[53] PARK C,PARK C,SHIN K,et al. Production of D-psicose from d-fructose by whole recombinant cells with high-level expression of D-psicose 3-epimerase from Agrobacterium tumefaciens[J]. Journal of Bioscience and Bioengineering,2016,121(2):186- 190.
[54] CHEN J,ZHU Y,FU G,et al. High-level intra- and extra-cellular production of D-psicose 3-epimerase via a modified xylose-inducible expression system in Bacillus subtilis[J]. Journal of Industrial Microbiology & Biotechnology,2016,43(11): 1577-1591.
[55] LI Z,LI Y,DUAN S,et al. Bioconversion of D -glucose to D -psicose with immobilized D -xylose isomerase and D -psicose 3-epimerase on Saccharomyces cerevisiaespores[J]. Journal of Industrial Microbiology & Biotechnology,2015,42(8):1117- 1128.
[56] YANG P,ZHU X,ZHENG Z,et al. Cell regeneration and cyclic catalysis of engineered Kluyveromyces marxianus of a D -psicose-3-epimerase gene from Agrobacterium tumefaciens for D-allulose production[J]. World Journal of Microbiology & Biotechnology,2018,34(5):7-13.
[57] LI C,ZHANG C,LIN J,et al. Enzymatic fructose removal from D-psicose bioproduction model solution and the system modeling and simulation[J]. Journal of Chemical Technology & Biotechnology,2018,93(5):1249-1260.
[58] SONG Y,NGUYEN QA,WI SG,et al. Strategy for dual production of bioethanol and D-psicose as value-added products from cruciferous vegetable residue[J]. Bioresource Technology,2017,223:34-39.
[59] HE X,ZHOU X,YANG Z,et al. Cloning,expression and purification of D-tagatose 3-epimerase gene from Escherichia coli JM109[J]. Protein Expression & Purification,2015,114:77-81.
[60] LI C,LIN J,GUO Q,et al. D-psicose 3-epimerase secretory overexpression,immobilization,and D-psicose biotransformation, separation and crystallization[J]. Journal of Chemical Technology & Biotechnology,2018,93(2):350-357.







