

 English
English French
French Spanish
Spanish Russian
Russian Korean
Korean Japanese
JapaneseThe Growth Factor in Spirulina Cultivation
1 Preface
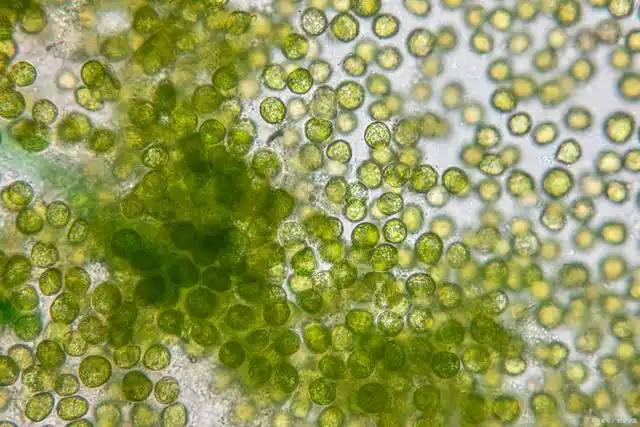
Spirulina, also known as Arthrospira, is a filamentous blue-green prokaryotic organism 7–400 µm long, 30–45 µm wide, and 7–10 µm in diameter (as shown in Figure 1). It mainly grows in tropical and subtropical regions and alkaline lakes. It has a living history of 3.5 billion years and is one of the oldest lifeforms on earth [1,2].
There are more than 50 species of spirulina, but only two species, Arthrospira maxima and Arthrospira platensis, are cultivated on a large scale internationally. Spirulina and its extracts are widely used in foods, feeds and cosmetics because of their rich nutrients, containing 60%–70% protein, 25% essential amino acids, 4% vitamins, etc., and various ingredients have certain biological activities, such as anti-oxidation and improving human immunity [3–5].
In recent years, many researchers have found through in vitro animal experiments that spirulina and its extracts have a series of biological functions on the human body, such as anti-oxidation, anti-inflammation, anti-tumor, immunity enhancement, and prevention of fatty liver [6−8], making spirulina have great potential in the biological and medical markets. At present, the world's spirulina consumption averages more than thousands of tons per year [9]. As spirulina's nutritional and health care functions have attracted more consumer attention, its market demand is also increasing year by year. However, due to the high cost of cultivating spirulina, the low yield per unit area, and the serious microbial contamination of the resulting product, improving spirulina cultivation technology to reduce costs and increase yield per unit area has become a research focus.

Spirulina has the characteristics of strong photosynthesis, fast growth and reproduction, strong environmental adaptability, and easy harvesting. It can be cultivated in closed and open pools in single batches or continuously, but its growth rate and component content are closely related to conditions such as nutrients, culture environment, and region. Generally, the spirulina culture solution can be harvested when the OD560 nm is 0.8 (approximately 0.34 g/L), but nutrient loss may occur during harvesting due to cell rupture [10]. Therefore, understanding the main influencing factors and their influence patterns in the cultivation of spirulina will allow the growth conditions of spirulina to be regulated according to the cultivation objectives (such as high yield, nutrient enrichment, etc.), thereby reasonably and effectively controlling the growth rate and component content of spirulina and obtaining high-quality and high-yield spirulina.
At present, there have been many reports on the biological functions of spirulina, but fewer and less in-depth reports on its cultivation conditions. For example, Tian Qiyang [11] studied the effect of nitrogen sources on spirulina growth. The results showed that ammonium sulfate at the same concentration could promote spirulina cell growth more than ammonium acetate, but the reason was not explained. Moreover, the study only focused on yield and did not consider the cost of cultivation. This work reviews the factors affecting spirulina growth and related issues such as cost and yield, providing theoretical guidance for the cultivation of spirulina with low cost, high yield and high quality.

2 Factors affecting spirulina growth
2.1 Nitrogen source
Protein is the main component of spirulina, accounting for 50% to 70% of its content. Nitrogen sources are the main raw materials for protein synthesis. The form and content of nitrogen sources in the culture medium have a significant effect on the growth and protein content of spirulina, and different nitrogen sources have different effects on the growth of spirulina. Currently, commonly used nitrogen sources include urea, sodium glutamate, NaNO3 and NH4Cl, ammonium acetate, etc. Table 1 shows the effect of different nitrogen sources and their concentrations on the final yield of spirulina. For example, 7.5 mmol/L urea and 15 mmol/L NaNO3 were used as nitrogen sources in a tubular photobioreactor, respectively, and single-batch culture was carried out for 8 d under the conditions of pH 9.5 and temperature 32 °C. the yields of spirulina cells were 486 and 515 mg/(L.d) respectively [12]. Although the urea concentration was half that of the NaNO3 concentration, the final yields of spirulina cultivated with the two were similar, indicating that the growth rate and yield of spirulina when using urea as a nitrogen source were higher. When the urea concentration exceeded 200 mg/L, the growth rate gradually decreased, mainly because [17,18]:
(1) each urea molecule provides 2 nitrogen atoms, while NaNO3 only supplies 1 nitrogen atom; (2) under alkaline conditions, urea can spontaneously hydrolyze in the culture solution to ammonia, which is easily absorbed by spirulina; (3) spirulina's absorption of nitrate first requires it to be reduced to nitrite, and then converted to ammonia under the action of enzymes. In addition, the reduction process consumes energy; (4) when the concentration of the nitrogen source is too high, excess nitrogen can be converted into toxic ammonium ions that inhibit cell growth. Sodium glutamate at a concentration of less than 1 g/L in the culture medium can increase the growth rate of spirulina as well as the content of protein and β-carotene, but at a concentration of more than 1 g/L, the growth rate decreases significantly due to limited metabolic capacity [11].
In recent years, NH4HCO3 has been increasingly used as a nitrogen source for spirulina cultivation for the following reasons: (1) it is less expensive, as the price of NH4HCO3 is lower than that of urea and NaNO3; (2) it can adjust the pH value and carbon source of the culture solution; and (3) the ammonia produced can inhibit the growth and reproduction of rotifers. Bao Yilu et al. [13] cultivated spirulina in a single batch, and the appropriate concentration of NH4HCO3 in the culture medium was 5.0 mmol/L, and the final spirulina yield could reach 260 mg/L. However, when the concentration of NH4HCO3 exceeded 5.0 mmol/L, the growth and reproduction of spirulina was inhibited, and it even disintegrated and died. Therefore, the final yield of spirulina can be effectively increased by combining different nitrogen sources during the cultivation process.
2.2 Carbon source
Spirulina is a photosynthetic microorganism that can use CO2 as a carbon source for photosynthesis under light conditions to produce energy and carbon-containing compounds. It can also synthesize carbohydrates (such as linolenic acid and polysaccharides) using organic or inorganic carbon sources in the absence of light. At present, simple organic compounds such as glucose, lactose and ethanol are commonly used as carbon sources for spirulina cultivation, mainly because they are easy to control and can obtain high yields of spirulina in a relatively short period of time. The effects of different carbon sources on the growth of spirulina and its component content vary greatly. For example, under the culture conditions of an illumination intensity of 5.0 klx and a carbon source concentration of 1 g/L, a single batch culture was carried out using ethanol, glucose and acetic acid as carbon sources in single batch culture for 14 days under the culture conditions of an illumination intensity of 5.0 klx and a carbon source concentration of 1 g/L, the final cell densities of spirulina were 2.17, 2.05 and 1.95 g/L, respectively, which were 1.2 to 1.4 times the cell density (1.57 g/L) of photoautotrophic culture with CO2 as the carbon source.
The content of linolenic acid (GLA) in total fatty acids in Spirulina is 23.6% (glucose), 24.2% (acetic acid), and 24.5% (ethanol) when using different carbon sources. Ethanol and acetic acid produce more GLA than glucose as a carbon source. The main reasons are: (1) Spirulina can directly convert acetic acid or ethanol into acetyl coenzyme A, a precursor of fat formation[19]; (2) organic by-products of glucose metabolism may inhibit the growth and reproduction of Spirulina at certain concentrations[14]. i.e. acetyl coenzyme A [19]; (2) organic by-products of glucose metabolism may inhibit the growth and reproduction of spirulina at certain concentrations [14].
The growth rate of spirulina is also affected by different concentrations of the same carbon source. For example, Tian Hua et al. [20] showed that when spirulina was cultivated with a mixed nutrient medium in Zarrouk medium and 4 klx light, the optimal concentration of glucose as a carbon source was 3 g/L. After 9 days, the dry weight of spirulina was 1.29 times that of the control group (without glucose). However, when the glucose concentration exceeds 4 g/L, the growth rate of spirulina is limited, mainly because: (1) glucose can activate the activity of spirulina cell physiological metabolic enzymes, enhance spirulina photosynthesis and increase the light saturation value, thereby producing higher spirulina biomass; (2) the control group spirulina is photosynthetic autotrophic, and HCO3− is its main carbon source. reaction HCO3−→CO32−+OH− causes a large change in the pH of the culture solution, which leads to inhibition of the growth rate of spirulina. However, it is worth noting that in an open industrial spirulina farming system, there is a complex competition between bacteria and algae. Adding an organic carbon source may disrupt the established bacteria-algae balance, and further research is needed.
Global warming in recent years is mainly caused by greenhouse gases such as CO2 produced by the combustion of fossil fuels. Cultivating photosynthetic microorganisms with CO2 as a carbon source to reduce CO2 emissions in the air has attracted much attention [21]. Ogbonda et al. [22] cultivated in a stepped spiral photobioreactor cultivated with a CO2 concentration of 0.44 g/(L.d) and a light intensity of 60 μmol/(m2.s) for 50 days in a stepped helix photobioreactor. The concentration of spirulina cells reached a maximum of 12.8 g/L, and the CO2 consumption rate exceeded 90% during the period from the 20th to the 50th day. However, excessive CO2 also had a certain inhibitory effect on spirulina growth. HCO3− is the main form of CO2 in aqueous solution and an important inorganic carbon source for spirulina cultivation. The CO2 concentration can be effectively controlled by adjusting the pH of the culture solution, thereby controlling the growth rate and final yield of spirulina [21]. Therefore, CO2 and other organic carbon can be reasonably combined as a carbon source for spirulina cultivation, which not only can obtain a large yield, but also reduce the emission of CO2 in the air.
2.3 pH and temperature
pH and temperature have a significant effect on spirulina cultivation. The growth rate of spirulina changes parabolically with pH and temperature. Spirulina can obtain the fastest growth rate and the highest protein content under the conditions of a culture solution with a pH of 9 to 10 and a temperature of 30 °C [1,22]. In addition to affecting the growth and reproduction of spirulina and cell morphology, pH and temperature also affect the content and activity of spirulina's active ingredients. Ismaielab et al. [23] found that the content of phycobiliproteins, chlorophyll and carotene in spirulina was highest when the pH of the culture medium was 8.5, reaching 91, 10.6 and 2.4 mg/g respectively; the antioxidant activity of spirulina was also highest when the pH was 9.0, its free radical scavenging capacity, reducing power and chelating capacity were 567%, 250% and 206% of the control group (2.5 μg BHT), respectively.
During the cultivation of spirulina, the consumption of HCO3− and nutrients and the production of metabolites caused the pH of the culture solution to rise and the temperature to increase. The production of free ammonium in the culture solution was the main reason for the decrease in spirulina production. Therefore, in a culture medium using NH4HCO3 as a nitrogen and carbon source, the pH value plays a key role in controlling the supply of nutrients to the spirulina. During the cultivation process, the pH value and temperature changes in the culture medium should be slowed down as much as possible to reduce the inhibitory effect on spirulina growth.
2.4 Light source
The light source is the main source of energy for the growth and reproduction of photosynthetic microorganisms. Spirulina is a photosynthetic microorganism, so the light source is also one of the most important factors affecting the growth and reproduction of spirulina. The wavelength, light duration and light intensity of the light source have a great influence on the growth rate and component content of spirulina. The final yield of spirulina increases with the increase of light intensity. If glucose is used as the carbon source, the final yield of spirulina is 1.24, 1.61 and 1.95 g/L when cultivated under light intensities of 2.0, 3.5 and 5.0 klx for 14 days, respectively [19]. Shi et al. [1] showed that when cultivated in a transparent glass tank, the growth rate of spirulina gradually increased with the increase of light wavelength, and the optimal light source wavelength was red light, and the optimal light time was 8 h/d. After 18 d of cultivation under red light (620–630 nm), blue light (465–475 nm) and green light (522–532 nm), the final yields of spirulina were 1.35, 1.18 and 1.08 g/L, respectively. The final yield of spirulina cultivated under red light increased by 56.69% compared to the control group (no light).
The light time also had a significant effect on the final yield of spirulina. When irradiated for 0–12 h per day for 18 d, the spirulina yield gradually increased when the light time was less than 8 h/d, after which there is no significant difference. The maximum yield of 1.44 g/L is obtained when the light is on for 8 h/d, which is 67.64% higher than the control group. The light source has a significant effect on the content of spirulina ingredients. When cultivated under ultraviolet light for 0–9 h, the fat content of spirulina increased by 29.5% compared to the control group (no ultraviolet light). Vonshak et al. [25] reported that when the light intensity exceeds the saturation light intensity of spirulina, photoinhibition occurs. Xia Jianrong et al. [26] showed that spirulina cultivated in high concentrations of CO2 can improve tolerance to high light intensity and reduce photoinhibition. Therefore, during the cultivation of spirulina in indoor containers or outdoor ponds, by continuously harvesting spirulina and adjusting the CO2 concentration, the OD650 nm value in the culture solution can be controlled within a certain range to ensure the light intensity in the culture solution, thereby obtaining the maximum growth rate of spirulina.
2.5 Rotifer contamination
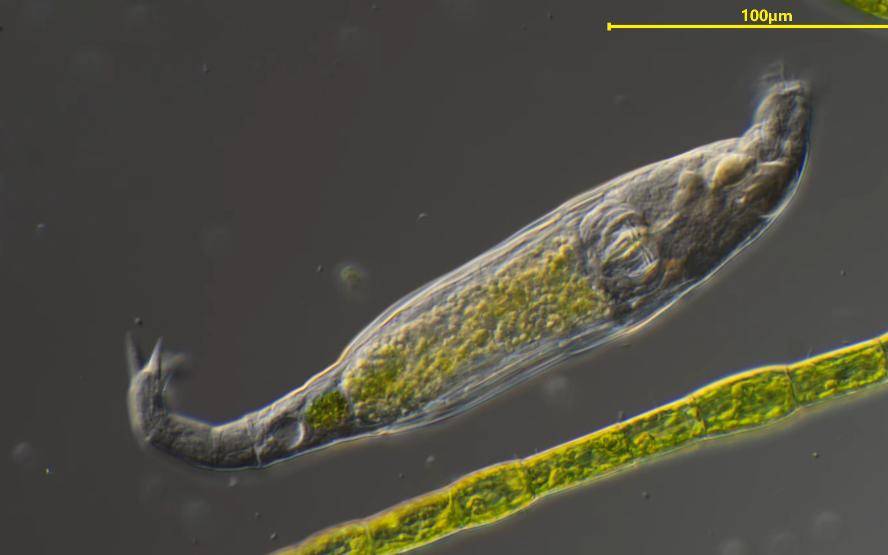
Rotifers are protozoa that feed exclusively on algae. They have a strong ability to adapt to their environment and reproduce both asexually and sexually. They are a serious hazard to spirulina cultivation. When the contamination by rotifers is extremely serious, it can cause a rapid decline in spirulina production or even a complete loss within 1 day. At present, there are two main methods used to reduce the contamination of algae culture by rotifers: (1) physical prevention methods, including disinfection, microporous filtration, etc., but the inhibitory effect is relatively poor; (2) chemical control methods, using chemicals (toluene, xylene, urea, etc.) to inhibit the growth and reproduction of rotifers, which has a better inhibitory effect. Since the artificial synthesis of chemical substances poses certain safety risks to commercial algae cultivation, natural inhibitors have become the main focus of current research [27-29].
For example, Huang et al. [28] found that matrine, matrine and matrine have a strong toxic effect on the ciliate Tetrahymena thermophila, and are relatively low in cost (about 10 yuan/g for matrine). The 24-hour LC50 values are 0.175, 0.061 and 2.13 μg/L, respectively. 1.76~2.13 μg/L matrine (a plant insecticide) can effectively reduce the number of rotifers and their reproductive capacity, and has no significant toxicity to the growth of spirulina cells and the photosynthesis of chlorophyll; 0.003~0.006 mg/L The combined solution (bitter vetch extract and kawain in a mass ratio of 1:9) can significantly inhibit the growth of rotifers within 3 d. At 9 d, all rotifers had died, and there was no significant effect on the final spirulina yield or its phycocyanin content [10]. An NH4HCO3 concentration of more than 200 mg/L in the culture medium has a significant inhibitory effect on the growth and reproduction of rotifers. After 24 hours, the rotifer population decreased from 52 to 6 per mL, and after 48 hours, its reproductive capacity was 0. The reason may be that the ammonia produced increased the extracellular glutamate content, resulting in acute central nervous system poisoning of the rotifers [28]. or the free ammonia may have a direct toxic effect on the rotifer cells.
2.6 Autotrophic growth inhibitors
Autotrophic growth inhibitors are metabolites produced during the cultivation of spirulina, such as macromolecular polysaccharides (molecular weight over 100 kDa), the remains of dead spirulina, and amines released by their decay. These have a serious inhibitory effect on the growth of spirulina, and their direct discharge can also cause environmental problems such as the death of aquatic plants and algal blooms.
Spirulina culture solution reused contains macromolecular polysaccharides, which can cause the growth rate of spirulina to decrease by 23% compared to fresh culture solution, nutrient salt absorption to decrease by more than 35%, protein content to decrease by 15%, and chlorophyll content to decrease by 35% [13]. The main reasons are as follows: (1) inhibitors can increase the hydrophobicity of the cell surface, leading to cell flocculation and sedimentation; (2) the effect of reducing chlorophyll content on photosynthesis; (3) reducing the activity of antioxidant system enzymes and nitrate reductase; (4) other microbial contamination [29]. In order to improve the reuse rate of the culture solution and reduce the environmental pollution caused by the culture solution, membrane separation technology and adsorbent adsorption are commonly used methods to remove organic matter from the spirulina culture solution. Yu et al. [30] found that the growth rate of spirulina in the culture medium filtered through a 100 kDa ultrafiltration membrane was 30% higher than that in the unfiltered culture medium, which was close to that in fresh culture medium. Wang et al. [31] used the macroporous adsorption resin S-8 to treat the culture medium, which reduced the extracellular polysaccharides by 62% and increased the growth rate of spirulina by 39.4%. Combined with the continuous harvesting of spirulina, it is imperative to develop technology for the separation and extraction of spirulina extracellular polysaccharides and extracellular proteins from spirulina culture broth.
2.7 Trace elements
Trace elements such as selenium, iodine, copper, zinc, and manganese also have a certain effect on the growth of spirulina [32]. Microalgae such as spirulina have a bio-concentrating effect on most metals. Low concentrations of metals have a certain promoting effect on the growth of spirulina, and spirulina with special uses can be grown. However, high concentrations of metals are toxic to spirulina by damaging the permeability of the cell membrane. Table 2 shows the concentrations of several trace elements that promote and inhibit the growth of spirulina. Selenium is an integral part of glutathione peroxidase, which is essential for human life and metabolism, and has the functions of maintaining normal cell metabolism and preventing and treating various diseases.
Organic selenium has lower toxicity and higher biological activity, and is an important source of selenium for the human body. Selenium in spirulina is mainly in the form of organic selenium (such as selenoprotein), so inorganic selenium can be converted to organic selenium through the enrichment effect of spirulina. Selenium concentrations below 0.1 mg/mL can promote spirulina growth, while also increasing the content of phycocyanin (C-PC) and carotene in spirulina [33]. However, selenium concentrations above 0.2 mg/mL are toxic and can directly lead to spirulina death. By adding selenium in small amounts, its cumulative concentration can reach 1 mg/mL, which can still promote the growth of spirulina. The selenium content in spirulina can reach up to 1.3 mg/g [39].
2.8 Other factors
The culture environment also has a certain effect on the growth of spirulina and the content of its components. Tolgal et al. [40] found that after 10 days of cultivation in a transparent incubator, the spirulina yield reached 0.99 g/L, but the protein content was low, at only 33.4%. When cultivated in polyethylene bags and ponds, the yield was only about 0.5 g/L, and the protein content was 54.5% and 58.3% respectively. The main reasons are: (1) in the transparent incubator, spirulina grows rapidly, nitrogen sources are quickly consumed, and protein synthesis is limited, resulting in a low protein content in spirulina; (2) the temperature change in a closed container is smaller than that in an outdoor pond, and it is easier to control, especially when cultivated in winter. The culture medium is an important nutrient for the growth and reproduction of spirulina. For example, Mg2+ promotes polysaccharide synthesis by activating enzyme activity, while K+ and Na+ maintain the normal osmotic pressure inside and outside the cell membrane.

Shi et al. [1] used a five-factor, four-level orthogonal experiment to obtain the composition and content of the optimal spirulina medium. The five main nutrients and their concentrations were NaHCO3 10 g/L, NaNO3 2 g/L, KH2PO4 0.6 g/L, MgSO4·7H2O 0.2 g/L and K2SO4 1.2 g/L. Vitamins also have a certain effect on the growth and content of spirulina. For example, Zhang Shaobin et al. [41] reported that 0.1–5 mg/L vitamin B5 can promote the growth of spirulina, with the greatest promoting effect at 0.5 mg/L. After 9 days of cultivation, the OD560 nm of the culture solution exceeded 0.9, and the content of phycocyanin reached a maximum of 0.02 mg/mL. Research [42] shows that the polyunsaturated fatty acids and cytochrome content of spirulina cultivated in seawater are significantly higher than those of spirulina cultivated in freshwater. A preliminary inference is that this is caused by the following reasons:
(1) seawater is rich in mineral nutrients; (2) seawater has good buffering properties and a slow pH change; (3) seawater has a high CO2 concentration. However, the specific reasons need to be further explored.
3 Summary and outlook
Spirulina powder is rich in nutrients and has certain biological activities. Coupled with the improvement of people's living standards and the strengthening of health awareness, the demand for it is gradually increasing. However, at present, spirulina companies are facing problems such as high cultivation costs, low yield per unit area, serious predation by rotifers, and serious microbial contamination of the resulting products, which has led to low spirulina production and limited its application. In addition, spirulina, as the ideal food of the 21st century, will become an integral part of future circular agriculture. Reducing production costs, increasing yields and controlling the risk of environmental pollution will become the main research directions for spirulina cultivation in the future.

With the rapid expansion of the population, the development of marine resources and the production of new marine foods have been put on the agenda, and there is an urgent need to improve the technology for cultivating spirulina using seawater. The type and concentration of nitrogen and carbon sources, CO2 concentration, light wavelength, and autotrophic growth inhibitors are parameters that have a significant impact on the growth rate of spirulina cells. Improving the cultivation technology of spirulina should start with the above parameters, and seek a spirulina cultivation technology with low cost and high yield. For example, during the indoor cultivation of spirulina, the growth rate of spirulina cells can be adjusted by controlling the type and concentration of nitrogen source, light wavelength and CO2 concentration, etc., while a certain amount of substances such as trichodermin can be added to inhibit the invasion of rotifers, reduce production costs, increase yields and reduce the risk of environmental pollution.
Reference:
[1] Shi W Q, Li S D, Li G R, et al. Investigation of Main Factors Affecting the Growth Rate of Spirulina [J]. Optik—International Journal for Light and Electron Optics, 2016, 127(16): 6688-6694, DOI: 10.1016/j.ijleo.2016.04.125.
[2] Vonshak A, Richmod A. Mass Production of the Blue-green Alga Spirulina: An Overview [J]. Biomass, 1988, 15(4): 233-247, DOI:10.1016/0144-4565(88)90059-5.
[3] Wang Z J, Zhang X W. Inhibitory Effects of Small Molecular Peptides from Spirulina (Arthrospira) platensis on Cancer Cell Growth [J]. Food Funct., 2016, 7(2): 781-788, DOI: 10.1039/C5FO01186H.
[4] Ozyurt G, Uslu L, Yuvka I, et al. Evaluation of the Cooking Quality Characteristics of Pasta Enriched with Spirulina platensis [J]. J.Food Qual., 2015, 38(4): 268-272, DOI: 10.1111/Jfq.12142.
[5] Jafari S M A, Rabbani M, Emtyazjoo M, et al. Effect of Dietary Spirulina platensis on Fatty Acid Composition of Rainbow Trout (Oncorhynchus mykiss) Fillet [J]. Aquaculture International, 2014, 22(4): 1307-1315
[6] Wu Q H, Liu L, Miron A, et al. The Antioxidant, Immunomodulatory, and Anti-inflammatory Activities ofSpirulina: An Overview [J]. Arch. Toxicol., 2016, 90: 1817-1840, DOI: 10.1007/S00204-016-1744-5.
[7] Yu J, Hu Y L, Xue M X, et al. Purification and Identification of Antioxidant Peptides from Enzymatic Hydrolysate of Spirulina platensis [J]. J. Microbiol. Biotechnol., 2016, 26(7): 1216-1223, DOI: 10.4014/Jmb.1601.01033.
[8] Ali H E A, Shanab S M, Abo-State M A, et al. Evaluation of Antioxidants, Pigments and Secondary Metabolites Contents in Spirulina platensis [J]. Appl. Mech. Mater., 2014, 625: 160-163.
[9] Khan Z, Bhadouria P, Bisen P S. Nutritional and Therapeutic Potential of Spirulina [J]. Curr. Pharm. Biotechnol., 2005, 6(5): 373-379.
[10] Huang Y, Liu J G, Wang H Y, et al. Treatment Potential of a Synergistic Botanical Pesticide Combination for Rotifer Extermination during Outdoor Mass Cultivation of Spirulina platensis [J]. Algal Research, 2014, 6: 139-144, DOI: 10.1016/J.Algal.2014.11.003.
[11] Tian Q Y. Advance in Effect of Factors on Growth of Spirulina platensis [J]. Animal Husbandry and Feed Science, 2009, (6): 113-115, DOI: 10.3969/J.Issn.1672-5190.2009.06.065
[12] Morocho-Jacome A L, Sato S, Capurro L D, et al. Simultaneous Use of Sodium Nitrate and Urea as Nitrogen Sources Improves Biomass Composition of Arthrospira platensis Cultivated in a Tubular Photobioreactor [J]. Eng. Life Sci., 2016. 16(4): 338-347, DOI: 10.1002/Elsc.201500051.
[13]Bao Y L, Liu M, Cong W, et al. Effect of Ammonium Bicarbonate as Nitrogen Source on Spirulina platensis Cultivation [J]. Food Science, 2012, 33(9): 193-196.
[14] Rodrigues M S, Ferreiral S, Converti A. Fed-batch Cultivation of Arthrospira (Spirulina) platensis: Potassium Nitrate and Ammonium Chloride as Simultaneous Nitrogen Sources[J]. Bioresour.Technol., 2010, 101(12): 4491-4498
[15] Jacome A L M, Converti A, Sato S, et al. Kinetic and Thermodynamic Investigation of Arthrospira (Spirulina) platensis Fed-batch Cultivation in a Tubular Photobioreactor Using Urea as Nitrogen Source [J]. J. Chem. Technol. Biotechnol., 2012, 87(11): 1574-1582, DOI: 10.1002/Jctb.3795.
[16] Cruz-Martinez C, Jesus C K C, Matsudo M C, et al. Growth and Composition of Arthrospira (Spirulina) platensis in a Tubular Photobioreaction Using Ammonium Nitrate as the Nitrogen Source in a Fed-batch Process [J]. Braz. J. Chem. Eng., 2015, 32(2): 347-356, DOI: 10.1590/0104-6632.20150322s00003062.
[17] Bezerrar P, Matsudo M C, Sato S, et al. Effects of Photobioreactor Configuration, Nitrogen Source and Light Intensity on the Fed-batch Cultivation of Arthrospira (Spirulina) platensis: Bioenergetic Aspects [J]. Biomass Bioenergy, 2012, 37: 309-317
[18] Yu S M, He L, Wu X, et al. Effect of Composition of Spirulina platensis Reused Culture Medium on the Growth of Its Cells [J]. Chin. J. Process Eng., 2016, 16(1): 151-155.
[19] Golmakani M, Rezaei K, Mazidi S, et al. r-Linolenic Acid Production by Arthrospira platensis Using Different Carbon Sources [J]. Eur. J. Lipid Sci. Technol., 2012, 114(3): 306-314, DOI: 10.1002/Ejlt.201100264.
[20] Tian H, Yu F, Wang J P. Influence of Glucose on the Culture of Spirulina platensis and Mechanism Analysis [J]. Food Research and Development, 2012, 33(7): 161-163, DOI: 0.3969/J.Issn.1005-6521.2012.07.046.
[21] Chen C Y, Kao P C, Tan C H, et al. Using an Innovative pH-stat CO2 Feeding Strategy to Enhance Cell Rowth and C-Phycocyanin Production from Spirulina platensis [J]. Biochem. Eng. J., 2016, 112: 78-85, DOI: 10.1016/J.Bej.2016.04.009.
[22] Ogbonda K H, Aminigo R E, Rebecca E, et al. Influence of Temperature and pH on Biomass Production and Protein Biosynthesis in a Putative Spirulina sp. [J]. Bioresour. Technol., 2007, 98(11): 2207-2211, DOI: 10.1016/J.Biortech.2006.08.028.
[23] Ismaielab M M S, Ei-Ayoutyb Y M, Pierycey-Normorea M. Role of pH on Antioxidants Production by Spirulina (Arthrospira) platensis [J]. Braz. J. Microbiol., 2016, 47(2): 298-304, DOI: 10.1016/J.Bjm.2016.01.003.
[24] Casazza A A, Ferrari P F, Aliakbrian B, et al. Effect of UV Radiation or Titanium Dioxide on Polyphenol and Lipid Contents of Arthrospira (Spirulina) platensis [J]. Algal Research, 2015, 12: 308-315, DOI: 10.1016/J.Algal.2015.09.012.
[25] Vonshak A, Guy R. Photoadaptation, Photoinhibition and Productivity in the Blue-green Alga, Spirulina platensis Grown Outdoors [J]. Plant, Cell & Environment, 1992, 15(5): 613-616, DOI: 10.1111/J.1365-3040.1992.Tb01496.X.
[26]Xia J R, Gao K S. Studies on Photoinhibition of Spirulina maxima Grown in High CO2 Concentration [J]. Acta Hydrobiologica Sinica, 2002, 26(1): 14-18, DOI: 10.3321/J.Issn:1000-3207.2002.01.003.
[27] Ferrando M D, Andreu-Moiner A. Acute Toxicity of Toluene, Hexane, Xylene, and Benzene to the Rotifers Brachionus Calyciflorus and Brachionus Plicatilis [J]. Bull. Environ. Contam. Toxicol., 1992, 49(2): 266-271.
[28] Huang Y, Li L, Liu J G, et al. Botanical Pesticides as Potential Rotifer-control Agents in Microalgal Mass Culture [J]. Algal Research, 2014, 4: 62-69, DOI: 10.1016/J.Algal.2013.08.001.
[29] Sun Y Y, Wang C H. Antialgal Mechanism of Growth-inhibitor Isolated from Marine Micoralga of Isochrysis Galbana [J]. Journal of Zhejiang University (Agriculture & Life Science), 2009, 35(1): 51-57, DOI: 10.3785/J.Issn.1008-9209.2009.01.007.
[30]Yu S M, Wu X, Wang J M, et al. Removing Technology Optimization of Auto-growth Inhibitors of Spirulina platensis by Ultra Filtration Membrane [J]. Journal of Henan University (Natural Science),2016,46(1): 69-75
[31]Wang L M, Liu M, Xue S Z, et al. Removal of Auto-growth Inhibitors of Spirulina platensis and Model Analysis [J]. Chin. J. Process Eng., 2012, 12(3): 478-483.
[32] Cone G, Lehmann B, Dussap C, et al. Uptake of Macrominerals and Trace Elements by the Cyanobacterium Spirulina platensis (Arthrospira platensis PCC 8005) under Photoautotrophic Conditions: Culture Medium Optimization [J]. Biotechnol. Bioeng., 2003, 81(5): 588-593, DOI: 10.1002/Bit.10504.
[33]Cao S Y, Liu H, Zhang S B. Advances in the Influence of Heavy Metals on the Growth of Spirulina [J]. Food Research and Development, 2011, 32(6): 171-172.
[34]Li Y Q,Gong L M, Cai Y, et al. Effects of Several Heavy Metals on the Growth of Spirulina platensis [J]. Light Industry Sciences and Technology, 2015, (2): 97-98.
[35]Zhang S B, Mu Y, Liu H, et al. Effect of Heavy Metal Chromium( Ⅵ ) on Growth of Spirulina [J]. Acta Agriculture Jiangxi, 2012, (2):145-146
[36]Li Y Y, Zhao N, Li S C, et al. Pb2+ Impacts on Growth of Two Spirulina Strains [J]. Journal of Biology, 2013, 30(4): 37-41, DOI: 10.3969/J.Issn.2095-1736.2013. 04.037.
[37]Li R Q, Wang C H. Study on Bioenrichment for Five Elements by Spirulina platensis [J]. Journal of Shanxi University, 2001, (2): 167-169
[38] Tian H, Wang JP, Zhang Y M.Effect of Iodine-enriched Cultivation and Current Research of Spirulina platensis [J]. China Brewing, 2011, (8): 16-18
[39]Chen T F,Cui X F,Yang F,et al. Culture of High Selenium-enriched Spirulina platensis with a Stepwise Selenium Addition Method and Its Effects on the Photosynthetic Pigment and Protein Contents of the Microalgae [J]. Food and Fermentation Industries, 2005, 31(8):48-51
[40] Tolgal G, Aysegu Z, Ak L. The Growth of Spirulina platensis in Different Culture Systems under Greenhouse Condition [J]. Turkish Journal of Biology, 2007, 31(1): 47-52.
[41]Zhang S B, Wang L, Liu G, et al. Effects of Vitamin B5 on the Growth of Spirulina and the Content of Algal Protein [J]. ChinaFeed, 2013, (13):18-19
[42]Xiang W Z, Li T, Wu H L, et al. The Strategic Studies on Developing Industry of Seawater Spirulina with Efforts[J]. Guangxi Sciences,2014,(6): 573-579







